MHC基因结构
主要组织相容性复合体(MHC)的完整概念是指脊椎动物某一染色体上编码主要组织相容性抗原、控制细胞间相互识别、调节免疫应答的一组紧密连锁基因群。人类MHC,即人的HLA基因位于第6号染色体上,称为HLA复合体。
编码HLA的HLA复合体位于人第6号染色体的短臂上。该区DNA片段长度约3.5~4.0 Kbp(千个碱基对),占人体整个基因组的1/3000。下图1显示HLA复合体结构。HLA复合体共有数十个座,传统上按其产物的结构、表达方式、组织分布与功能可将这些基因座分为三类(本站“编码HLA基因的分类及其命名”部分已经做了详细描述,此处就作简要介绍)。
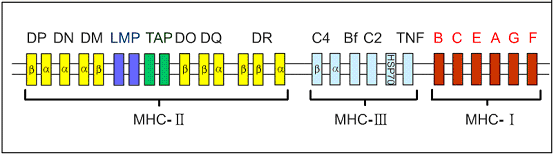
图1 位于第6号染色体上的HLA复合体结构示意图
1. MHC-Ⅰ类基因
在Ⅰ基因区内存在多达31个有关的Ⅰ类基因座,其中HLA-A、HLA-B和HLA-C为经典的HLA-Ⅰ类基因,其它基因的产物分布有限,且其功能不明,另外还有许多假基因。
2. MHC-Ⅱ类基因
HLA-Ⅱ类基因区包括近30个基因座,其中经典的Ⅱ类基因一般指DR、DP和DQ,它们编码的产物均为双肽链(α、β)分子。近年来,陆续发现了一些位于Ⅱ类基因区的新基因座,其中某些基因的产物与内源性抗原的处理与呈递有关。
3. MHC-Ⅲ类基因
HLA-Ⅲ类基因区域至少已发现36个基因座,其中C2、C4、Bf座编码相应的补体成分,另外还有21羧化酶基因(CYP21A、B)肿瘤坏死因子基因(TNF A、B)以及热休克蛋白70(heat shock protein70,HSP70)基因。补体C4由二个不同的基因(C4A与C4B)编码。
HLA抗原的分子结构
1987年Bjorkman等首先借助X线晶体衍射技术弄清了HLA-A2分子的立体结构。其后, HLA-Ⅰ、Ⅱ类分子结构的研究也取得了进展,从而对这些分子的生物学功能提供了较确切的解释。下面就以HLA-Ⅰ、Ⅱ类抗原为例介绍HLA抗原的分子结构。
(一)HLA-Ⅰ类分子
所有的HLA-Ⅰ类分子均含有二条分离的多肽链,一条是由MHC基因编码的α链或称重链(44kD)。根据对HLA-A2和Aw68分子的晶体结构分析,Ⅰ类分子可分为四个区:(图2):
① 氨基端胞外多肽结合区:
该区由二个相似的各包括90个氨基酸残基的片段组成,分别称为α1和α2。该功能区含有与抗原结合的部位。后者呈深槽状,其大小与形状适合于已处理的抗原片段,约容纳8~10个氨基酸残基。Ⅰ类分子的多态性残基也位于该区域。
② 胞外Ig样区:
该区又称为重链的α3片段,包括90个氨基酸残基,与免疫球蛋白的恒定区具有同源性。Ⅰ类分子与TC细胞表面CD8分子的结合部位即在α3片段。Ⅰ类分子的β链又称β2微球蛋白,也结合于该区。β链由第15号染色体的基因编码,它不插入细胞膜而游离于细胞外。β2微球蛋白与α1、α2、α3片段的相互作用对维持Ⅰ类分子于然构型的稳定性及其分子表达有重要意义。
③ 跨膜区:
该区氨基酸残基形成螺旋状穿过浆膜的脂质双层,将类分子锚定在膜上。
④ 胞浆区:
该区位于胞浆中,可能与细胞内外信息传递有关。
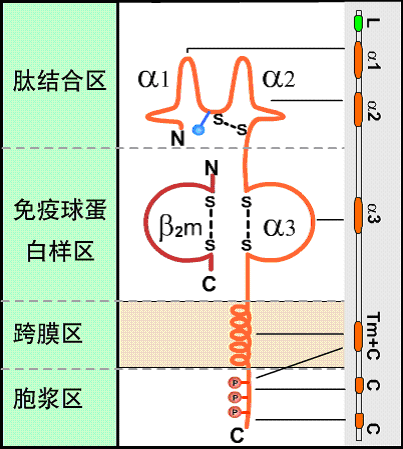
图2 HLA-Ⅰ类分子结构示意图
(二)HLA-Ⅱ类分子
所有的Ⅱ类分子均由二条以非共价键连接的多肽链(α、β)组成。二条链的基本结构相似,但分别由不同的MHC基因编码,且均具有多态性。虽然Ⅱ类分子的晶体衍射结构尚未得到,但光谱分析已证明与Ⅰ类分子具有某种相似性。Ⅱ类分子二条多肽链也可分为四个区,见(图3):
①肽结合区:
α链与β链的胞外部位均可再分为二个各含90个氨基酸残基的片段,分别称为α1、α2和β1、β2。肽结合区包括α1和β1片段,该二片段构成肽结合的裂隙(cleft),约可容纳14个氨基酸残基。Ⅱ类分子的多肽残基主要集中在α1和β1片段,这种多态性决定了多肽结合部位的生化结构,也决定了与肽类结合以及T细胞识别的特异性和亲和力。
②Ig样区:
此区由α2和β2片段组成,两者均含链内二硫键,并属于Ig基因超家族。在抗原呈递过程中,TH细胞的CD4分子与Ⅱ类分子结合的部位即位于该Ig样非多肽态区域。
③跨膜区和胞浆区:
该二区与Ⅰ类分子α链的相应区域结构相似。
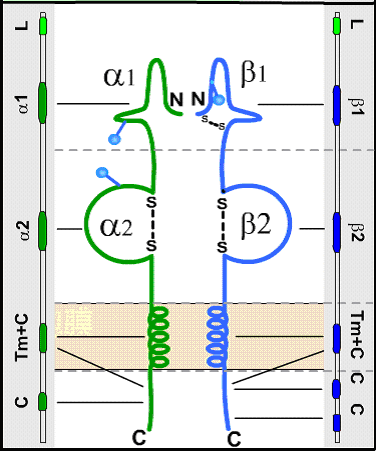
图3 HLA-Ⅱ类分子结构示意图
编码HLA基因复合体(MHC)的遗传特征
HLA复合体具备某些有别于其它真核基因系统的特征。
1. 连锁不平衡
HLA复合体各等位基因均有其各自的基因频率。基因频率是指某一特定等位基因占该基因座中全部等位基因总和的比例。随机婚配的群体中,在无新的突变和自然选择的情况下,基因频率可以代代维持不变。一个HLA位点的等位基因与另一个或几个位点的等位基因在某一单倍型出现的频率应等于各自频率的乘积。然而在很多情况下,预期的单倍型频率往往与实际检测的频率相差很大,在不同的地区或不同的人群,某些基因相伴出现的频率特别高,这种现象称为连锁不平衡(linkage disequilibrium)。例如,在北欧白人中HLA-A1和HLA-B8频率分别为0.17和0.11。若随机组合,则单体型A1-B8的预期频率为0.17×0.11=0.019。但实际所测行的A1-B8单体型频率是0.088故A1-B8处于连锁不平衡,实测频率与预期频率间的差值(△0.088-0.19=0.069)为连锁不平衡参数。在HLA复合体中已发现有50对以上等位基因显示连锁不平衡。HLA基因连锁不平衡的发生机制目前尚不清楚,但已经发现某些疾病的发生与HLA复合体中某些特定的等位基因密切相关;某些连锁不平衡倾向于出现在某些区域、某些人种和某些民族。深入探讨连锁不平衡的发生机制无疑将有助于对某些疾病的诊断和治疗,亦将为人类学研究增添新的内容。
2. 单体型遗传方式
HLA复合体是一组紧密连锁的基因群。这些连锁在一条染色体上的等位基因很少发生同源染色体间的交换,构成一个单体型(haplotype)。在遗传过程中,HLA单体型作为一个完整的遗传单位由亲代传给子代。有必要区分HLA表型、基因型与单体型这三个概念。某一个体HLA抗原特异性型别称为表型(phenotype);HLA基因在体细胞两条染色体上的组合称为基因型(genotype);HLA基因在同一条染色体上的组合称为单体型(haplotype)(表1)。
表1 HLA表型、基因型与单体型的区别
受检者 甲 乙 丙
A1 A2B8 B12 A1 A1B8 B12 A1 A1B8 B8
表型 HLA-A1、2:B8、12 HLA-A1:B8、12 HLA-A1、B8
基因型 HLA-A1、A2HLA-B8、B12 HLA-A1、A1HLA-B8、B12 HLA-A1、A1HLA-B8、B8
单体型 HLA-A1、B8/A2、B12 HLA-A1、B8/A1、B12 HLA-A1、B8/A1、B8
二倍体(diploid)生物的每一细胞均有两个同源染色体组,分别来自父母双方。故子女的HLA单体型也是一个来自父方,一个来自母方。在同胞之间比较HLA单体型型别只会出现下列三种可能性:二个单体型完全相同或完全不同的机率各占25%;有一个单体型相同的机率占50%。至于亲代与子代之间则必然有一个单体型相同,也只能有一个单体型相同(图4)。这一遗传特点在器官移植供者的选择以及法医的亲子鉴定中得到了应用。
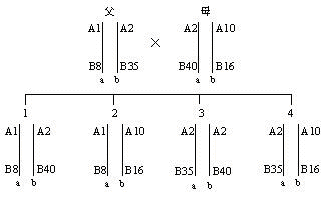
图4 HLA单体型遗传示意图
注:a、b、c、d代表单体型;A1、B8、A2、B35等代表HLA基因座等位基因
由于存在连锁不平衡, 某些单体型在群体中可呈现较高的频率, 并较之单一HLA基因型别更能显示人种和地理族(ethnic group)的特点。例如,中国汉族人中具有特征性的HLA单体型主要有A2-B46-Cw3-DR9-DQ9-Dw23和 A33-B17-Cw2-DR3-DQ2-Dw3。 检测单体型比分析单一的等位基因频率更有助于从无血缘关系人群中搜寻HLA相匹配的器官移植供者。然而, 连锁不平衡却不利于分析和寻找调控免疫应答或决定疾病易感性的关键性基因成份。举例说,如果DQ座位的等位基因HLA-DQB1*0301对某一疾病的发生起决定性作用,但此0301又和DR座位的DRB1*0901及B座位的B*4601呈现连锁不平衡,可能使人误认为 DRB1*0901或B*4601和疾病关联。实际上只有DQB1*0301属于该病的原发关联 (primary association)成分,其余两个位于DR和B座位上的等位基因产物属于次级关联。
3. 多态性现象
HLA复合体是迄今已知人体最复杂的基因复合体,有高度的多态性。多态性 (polymorphism) 指一个基因座位上存在多个等位基因 (allele)。对某一个基因座位,一个个体最多只能有两个等位基因,分别出现在来自父母方的同源染色体上。因而MHC的多态性是一个群体概念,指群体中不同个体在等位基因拥有状态上存在差别。多态性和多基因现象,是从不同水平对MHC的多样性进行描述:多基因性着重于同一个个体中MHC基因座位的变化;而多态性指群体中各座位等位基因的变化。据1999年的统计,整个HLA复合体等位基因总数已达到1031个,其中等位基因数量最多的座位是HLA-B(301个)和HLA-DRB1(227个)。
HLA的多态性程度可以从两方面进行估计:一是每个基因座位上的杂合体比例。按照种群遗传学的理论,每个座位上的纯合体比例等于基因频率的平方和。一般来讲等位基因越多,纯合体的比例也就越低。举例来说,在每个等位基因频率都相同的情况下,在双等位基因系统中,纯合体占50%;在4个等位基因的系统中,纯合体占25%;在10个等位基因的系统中,纯合体仅占10%。衡量HLA多态性的另一个标志是表现型的数目,ABO血型系统的表现型有4种;Rh系统有18种;而HLA系统的表现型则多达上亿种。
对HLA系统的不同基因座位,和同一座位不同等位基因的命名原则是,星号(*)前为基因座位,星号后为等位基因。例如HLA-A*0103代表HLAⅠ类基因A座位的第103号等位基因;HLA -DRB1*1102代表Ⅱ类基因DRB1座位第1102号等位基因。1102并不表明DRB1座位已发现一千一百多个等位基因,而是根据等位基因的结构,通常再分成若干主型 (generic type), 1102是DRB1座位第11主型第2号等位基因。同理,HLA A*0103是A座位第1主型第3号等位基因。
HLA的多态性现象乃由于下列原因所致:①复等位基因(multiple alleles):位于一对同源染色体上对应位置的一对基因称为等位基因(allele);由于群体中的突变,同一座的基因系列称为复等位基因。前已述及,HLA复合体的每一座均存在为数众多的复等位基因,这是HLA高度多态性的最主要原因。由于各个座位基因是随机组合的,故人群中的基因型可达108之多。②共显性(codominance):一对等位基因同为显性称为共显性。HLA复合体中每一个等位基因均为共显性,从而大大增加了人群中HLA表型的多样性,达到107数量级。因此,除了同卵双生外,无关个体间HLA型别全相同的可能性极小。
HLA基因结构的变异为进化中的选择提供了基础。变异的发生涉及基因突变、基因重组(同源染色体之间的交换)和基因转换等多种机制。基因转换 (gene conversion)指两条染色体非同源部分发生DNA片段的转移。基因转换可产生新的等位基因。其他真核细胞的基因较少发生基因转换,但在MHC多态性产生中却十分常见。 MHC变异的产生属于一组偶发事件,这些偶发的变异能否以新等位基因的形式被一代一代地保存下来,取决于自然选择。如果新出现的等位基因,在提呈病原体的关键性抗原肽方面有独到之处,则相应个体较易启动有效的免疫应答,显示较强的抗病能力和较低的死亡率。换言之,这一新等位基因的拥有者,会有较多的机会把该等位基因传递给后代。于是,在群体水平新的等位基因投向该物种基因库的比例将大于其它的基因,两者之差构成选择压力 (selection pressure)。选择压力使新基因频率逐渐上升。结果,新等位基因在群体中得到积累,形成多态性。
多态性主要为经典的Ⅰ类基因和Ⅱ类基因所有,这是与Ⅰ、Ⅱ类基因产物参与提呈抗原肽有关的。由于不同的MHC等位基因产物可以递呈结构不同的抗原肽,并诱发出特异性和强度不同的免疫应答,而这种MHC分子在结构和抗原递呈能力上的不同,往往表现在带有不同MHC等位基因的个体水平,这意味着,MHC多态性从基因的储备上,造就了对抗原(病原体)入侵反应性和易感性不同的个体。这一现象的群体效应,是赋于物种极大的应变能力,使之能对付多变的环境条件及各种病原体的侵袭。应该说,这是MHC高度多态性具有强大生命力的体现,是长期自然选择的结果。
总之,HLA的高度多态性显示了遗传背景的多样性,这可能是高等动物抵御不利环境因素的一种适应性表现,从而维持种属的生存与延续具有重要的生物意义,但也对组织移植过程中寻找配型合的供体带来很大的困难。
|





 沪ICP备10027510号-1
沪ICP备10027510号-1